Inhalt:
Es gibt nur wenige körperliche Merkmale und Eigenschaften, die wir nicht von unseren Vorfahren geerbt haben. Jeden Tag und bei jeder Tätigkeit stoßen wir auf eine Erbschaft, die uns die vorherige Generation durch unsere Anlagen – die Gene – weitergegeben hat. Es ist schon verblüffend, was wir alles „von Opa“, „von Oma“ oder auch „vom Onkel vierten Grades“ an uns haben. Und nicht anders verhält es sich bei allen lebenden Organismen, die uns umgeben – Blumen, Bäume, Insekten, Vögel und andere Tiere verfügen über eine Ansammlung von Anlagen, die über viele Generationen angehäuft wurden.
Mit der Erkenntnis der wechselseitigen Beziehungen zwischen den Generationen und der Art der Weitergabe der Erbinformationen beschäftigt sich eine komplexe und schöne Wissenschaft – die Genetik.
Sobald uns allerdings dieses Wort über die Lippen kommt, entsteht unter Züchtern Panik – und anschließend gibt es nur eine Reaktion: „Kommen Sie mir nicht mit so etwas Unverständlichem"!
Vor etlichen Jahren habe ich an der Hochschule Genetik studiert, habe darin meine Prüfung abgelegt, aber nachdem ich die Alma mater verlassen habe, war meine Einstellung zur Vererbungslehre beinahe genauso wie die anderer Hundehalter. Es war für mich nur ein fürchterliches und unanwendbares Schulfach! Aber im Unterschied zu anderen Kynologen hatte ich ein Riesenglück. Dank jugendlicher Unbekümmertheit habe ich einen bedeutenden tschechischen Kynologen, Herrn František Horák, kennengelernt, einen langjährigen Mitarbeiter am Physiologischen Institut der Tschechoslowakischen Akademie der Wissenschaften (ČSAV), Schöpfer zweier Rassen und Praktiker der angewandten Genetik mit großer Erfahrung. Mehr als zwanzig Jahre, bis zu seinem Tod, war ich seine dankbare Schülerin, und er hat mich gelehrt, die Genetik nicht als etwas Feindliches anzusehen, sondern als eine wirkliche Hilfe. Durch die Gespräche mit ihm habe ich die Scheu vor dieser Wissenschaft verloren, deren Kenntnis für jeden Züchter, der diesen Namen verdient, unabdingbar ist.
Ich möchte Ihnen gerne auf ein paar Seiten die elementaren Prinzipien der Vererbungslehre näherbringen und Ihnen vor allem ihre praktische Anwendbarkeit zeigen.
Ich gehe davon aus, daß viele von Ihnen noch nie etwas mit Genetik zu tun hatten bzw. nach dem ersten Kontakt davon wieder Abstand genommen haben. Wir bitten diejenigen, die sich professionell mit Genetik beschäftigen, um Nachsicht, wenn wir die neuesten Erkenntnisse der genetischen Forschung nicht berücksichtigt haben. Wir sehen es nicht als unsere Aufgabe an, unsere Mitglieder zu allseitig gebildeten Experten dieser komplizierten Wissenschaft zu erziehen, zu der die Genetik heute geworden ist – wir möchten nur auf möglichst populäre Weise die breite Masse der Züchter mit den Ursachen einiger Erscheinungen bekannt machen, auf die sie in der Praxis stoßen.
Beginnen wir also bei „Adam und Eva“.
 Die Fähigkeit, bestimmte Eigenschaften und Merkmale der Familie an die Nachkommen weiterzugeben, nennt man Vererbbarkeit oder genetische Veranlagung. Dagegen bezeichnet man die Fähigkeit, immer wieder aus lebendem Material
unterschiedliche Lebensformen zu bilden als Veränderlichkeit oder Variabilität.
Die Wissenschaft, die sich mit der Vererbbarkeit beschäftigt, heißt Genetik. Als selbständigen Zweig der Wissenschaft ist sie ziemlich spät entstanden und ihr Name geht auf einen Vorschlag von William Bateson im Jahre 1906
zurück. Die Priorität bei der Formulierung der genetischen Grundlagen gebührt dem Abt des Augustinerklosters zu Brünn Johann Gregor Mendel, der bereits 1865 die Ergebnisse seiner Kreuzungsexperimente mit verschiedenfarbigen Erbsen veröffentlichte. Er stellte dabei die wichtigen Gesetzmäßigkeiten fest, die wir heute als Mendelsche Regeln bezeichnen und die eine außerordentliche Bedeutung bei der Entwicklung der Genetik hatten.
Die Fähigkeit, bestimmte Eigenschaften und Merkmale der Familie an die Nachkommen weiterzugeben, nennt man Vererbbarkeit oder genetische Veranlagung. Dagegen bezeichnet man die Fähigkeit, immer wieder aus lebendem Material
unterschiedliche Lebensformen zu bilden als Veränderlichkeit oder Variabilität.
Die Wissenschaft, die sich mit der Vererbbarkeit beschäftigt, heißt Genetik. Als selbständigen Zweig der Wissenschaft ist sie ziemlich spät entstanden und ihr Name geht auf einen Vorschlag von William Bateson im Jahre 1906
zurück. Die Priorität bei der Formulierung der genetischen Grundlagen gebührt dem Abt des Augustinerklosters zu Brünn Johann Gregor Mendel, der bereits 1865 die Ergebnisse seiner Kreuzungsexperimente mit verschiedenfarbigen Erbsen veröffentlichte. Er stellte dabei die wichtigen Gesetzmäßigkeiten fest, die wir heute als Mendelsche Regeln bezeichnen und die eine außerordentliche Bedeutung bei der Entwicklung der Genetik hatten.
 Der elementare Baustein des Körpers ist die Zelle. Die Körperzelle besteht aus einem Kern und einer umgebenden Substanz, dem Protoplasma. Träger der Erbinformation sind sowohl der Zellkern (Vererbung durch den Zellkern) als auch die umgebende Substanz. Der Zellkern beinhaltet Chromosomen, das sind fadenförmige Gebilde, deren Form und Anzahl gattungsspezifisch sind. Die Chromosomen sind paarweise angeordnet. Es sind immer zwei Chromosomen gleich, nur bei einem Paar sind sie ungleich: beim Geschlechtschromosom, das das Geschlecht des neuen Lebewesens bestimmt. Die weiblichen Chromosomen bezeichnet man nach ihrer Form als X, die männlichen als Y. Ein weibliches Säugetier hat immer zwei X-Chromosomen, ein männliches ein X- und ein Y-Chromosom. Ein Hund besitzt 38 somatische (körperliche) Chromosomenpaare und ein Geschlechts-Chromosomenpaar. Der Wuchs des Organismus erfolgt durch Zellteilung, wobei sich auf komplexe Art alle Körperzellen verdoppeln und aus den Mutterzellen völlig gleiche Tochterzellen entstehen (auch bei den 39 Chromosomenpaaren beim Hund).
Der elementare Baustein des Körpers ist die Zelle. Die Körperzelle besteht aus einem Kern und einer umgebenden Substanz, dem Protoplasma. Träger der Erbinformation sind sowohl der Zellkern (Vererbung durch den Zellkern) als auch die umgebende Substanz. Der Zellkern beinhaltet Chromosomen, das sind fadenförmige Gebilde, deren Form und Anzahl gattungsspezifisch sind. Die Chromosomen sind paarweise angeordnet. Es sind immer zwei Chromosomen gleich, nur bei einem Paar sind sie ungleich: beim Geschlechtschromosom, das das Geschlecht des neuen Lebewesens bestimmt. Die weiblichen Chromosomen bezeichnet man nach ihrer Form als X, die männlichen als Y. Ein weibliches Säugetier hat immer zwei X-Chromosomen, ein männliches ein X- und ein Y-Chromosom. Ein Hund besitzt 38 somatische (körperliche) Chromosomenpaare und ein Geschlechts-Chromosomenpaar. Der Wuchs des Organismus erfolgt durch Zellteilung, wobei sich auf komplexe Art alle Körperzellen verdoppeln und aus den Mutterzellen völlig gleiche Tochterzellen entstehen (auch bei den 39 Chromosomenpaaren beim Hund).
Ein neues Lebewesen entsteht durch die Verbindung zweier Geschlechtszellen – Ei und Spermium. Weil es bei diesem Akt aber zu einer Verdoppelung der ursprünglichen Erbsubstanz und daher zur Entstehung eines Hybriden (Bastards) kommen könnte, verläuft die Entstehung der Geschlechtszellen völlig anders. Durch eine sog. Reifeteilung (Meiose) reduziert sich die Anzahl der Chromosomen in der Zelle auf die Hälfte, so daß Spermium und Ei nur einen einfachen (haploiden) Chromosomensatz enthalten. Der Nachkomme hat auf diese Weise wieder die normale Anzahl an Chromosomen, wobei er die eine Hälfte seiner Erbinformationen vom Vater und die andere Hälfte von der Mutter bekommen hat.
Die grundlegende Erbeinheit ist das Gen (Anlage). Die Gene sind in den Chromosomen hintereinander wie Perlen auf einer Schnur aufgereiht, und normalerweise hat jedes Gen seinen festen Platz. Unter dem Einfluß verschiedener Faktoren (z.B. Bestrahlung) kann es zu einer Veränderung des Originalgens kommen – einer Mutation des Gens. Derart veränderte Gene (Allele) belegen die gleiche Stelle in den Chromosomen wie die Originalgene.
Einige der körperlichen Merkmale – qualitative Merkmale – sind von sehr einfacher Art, man sagt, daß sie Gene von großer Wirkung zur Grundlage haben. Im wesentlichen handelt es sich um Merkmale und Eigenschaften vom Typ Entweder-Oder: Farbe, Ohrenform, mit oder ohne Hörner.
Sehr komplex ist dagegen die Erbanlage etwa von Charakter- und Gebrauchseigenschaften – quantitative Merkmale: Statur, Körpergewicht, Leistungsfähigkeit, Fruchtbarkeit. Sie haben zur Grundlage sogenannte Gene von geringer Wirkung, Polygene, die sich in ihrer Wirksamkeit gleichsam summieren.
Die Gesamtheit der Gene nennt man den Genotyp. Der Phänotyp ist dagegen das äußere Erscheinungsbild der ererbten Eigenschaften, Ergebnis der Auswirkungen der Lebensbedingungen auf die Erbanlagen des Tieres. Dies läßt sich am besten an einem Beispiel zeigen: die Farbe des Hundes ändert sich nicht durch die Lebensbedingungen (wenn man außer acht läßt, ob er sauber oder schmutzig ist), aber die Statur des Tieres und natürlich seine Leistungskraft kann man durch die Lebensbedingungen beeinflussen. Eine schlechte Ernährung änderts zwar nicht die Erbanlage, die der Welpen von den Eltern mitbekommt, beeinträchtigt aber sein äußeres Erscheinungsbild – der Welpe kann klein und schwach bleiben, kaum fruchtbar, ohne Interesse an irgendeiner körperlicher Aktivität.
Kommen wir zu Erklärung weiterer Begriffe.
Versuche haben ergeben, daß einige Gene bei ihrer Entwicklung anderer Gene oder auch ihre eigenen Allele beeinträchtigen – sie sind vorherrschend, dominant und man kennzeichnet sie mit großen Buchstaben. Unterdrückte, zurücktretende Gene bezeichnet man als rezessiv und kennzeichnet sie mit kleinen Buchstaben. Da der Nachkomme die eine Hälfte seines Genotyps von der Mutter und die andere Hälfte vom Vater bekommt, ist jedes Merkmal oder jede Eigenschaft immer in zwei Genen vorhanden. Sofern diese Gene von gleicher Qualität sind, sagt man, daß das Tier für dieses Merkmal reinerbig – homozygot ist, und zwar entweder dominant (beide Gene sind dominant AA) oder rezessiv (beide Genesindrezessiv aa). Bei ungleicher Qualität der Gene ist das Tier mischerbig, eine "Kreuzung" – heterozygot Aa. Jedes Tier ist für viele Merkmale heterozygot, ein absolut homozygotes Tier kann es nicht geben. Da die Erklärung von mehreren Merkmalen sehr kompliziert wäre, reden wir weiterhin nur über ein einzelnes Merkmal und gehen von der Annahme aus, das Elternpaar sei homozygot.
 Mendelsche Regeln
Mendelsche Regeln
1) Die Uniformitäts- oder Gleichheitsregel in F1 und ihre Gültigkeit auch bei reziproker Kreuzung.
Wir wollen es an einem Beispiel erklären: Wir vereinigen die Eltern AA und aa in der Eltern- bzw. Parentalgeneration P. Damit erwirken wir die erste Tochter bzw. Filialgeneration F. Aus den bisherigen Ausführungen wissen wir, daß die Geschlechtszellen (Gameten) nur einen einfachen Chromosomensatz enthalten, also für jedes Merkmal nur ein einziges Gen. Die Eltern produzieren die Gameten AAaa. Alle ihre Nachkommen in F1 sind nur Kombinationen des Gameten A mit dem Gamet a – also heterozygot Aa.
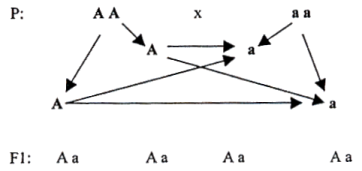
Mit reziproker Kreuzung meinen wir die zwei Kombinationsmöglichkeiten zwischen den Eltern:
1) Vater A A, Mutter a a
2) Vater a a, Mutter A A
Hinsichtlich der Merkmale, deren genetische Grundlage im Zellkern liegt (Vererbung durch den Zellkern) gilt, daß die Resultate bei reziproker Kreuzung in F1 gleich sind. Es ist beispielsweise völlig egal, ob wir einen schwarzen Vater und eine blaue Mutter nehmen oder einen blauen Vater und eine schwarze Mutter. Bei einigen Merkmalen dagegen ist die umgebende Substanz des Kerns der Träger der Erbinformation und da gilt diese Regel natürlich nicht. Ein Beispiel ungleicher Resultate reziproker Kreuzung ist etwa das unterschiedliche Ergebnis einer Verpaarung einer großen Hündin mit einem kleinen Rüden (die Welpen sind bei der Geburt groß nach der Mutter) und einer kleinen Hündin mit einem großen Rüden (die Welpen sind bei der Geburt klein nach der Mutter). Auch wenn die Erbinformation in den Geschlechtschromosomen liegt (geschlechtsgebunden ist), sind die Resultate reziproker Kreuzung unterschiedlich.
2) Die Spaltungsregel der Erbanlage in F2 sowie die Reinheit der Erbanlage.
Die Eltern Aa und Aa aus der ersten Filialgeneration bilden die Gameten AaAa, und daher können sich bei ihrer Verpaarung nicht nur die heterozygoten Nachkommen Aa entwickeln, sondern auch die homozygoten AA, aa. Es kommt zur Abspaltung des Genotyps Aa.
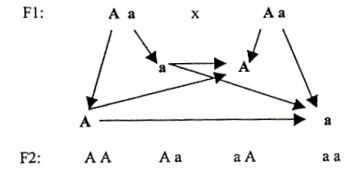
Einzelne Gene gelangen in den Gameten und damit auch in den Nachkommen in Reinform – also entweder A oder a, sie vermischen sich nicht miteinander, auch wenn das äußere Erscheinungsbild manchmal darauf hinzuweisen scheint. Bei einigen körperlichen Merkmalen (typischerweise Farbe oder Anzahl der Zähne) weisen die Gene eine sog. vollständige Dominanz auf, d.h.solange sie präsent sind, treten sie immer in Erscheinung. Dagegen kommt es bei anderen Merkmalen (Nasenlänge, Qualität des Fells) zu einer sog. unvollständigen Dominanz,nämlich dann, wenn sich das äußere Erscheinungsbild gleichsam vermischt. Es handelt sich aber niemals um eine Vermischung der Gene als solcher!
3) Die Regel der freien Kombinierbarkeit der Erbanlage.
J. G. Mendel fand heraus, daß die einzelnen Gene untereinander frei kombinierbar sind, sofern sie sich auf verschiedenen Chromosomen befinden.
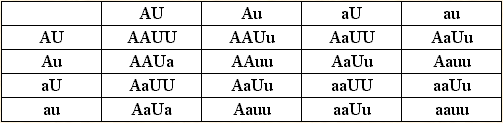
Verpaaren wir einen schwarzen Rüden mit Kippohren mit einer braunen Hündin mit Stehohren. Wir markieren ihren Genotyp: der Rüde AAUU, die Hündin aauu. Ihre Nachkommen werden in F1 AaUu sein – Welpen mit Kippohren. Die F1-Individuen bilden die Gameten AUAuaUau. Faßt man sie in einer Tabelle zusammen, zeigt sich, wie viele Kombinationsmöglichkeiten dieser zwei Gene es gibt.
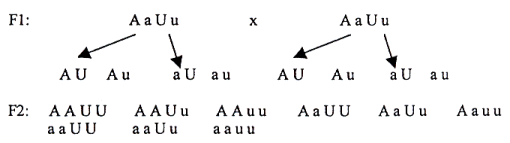
Tabelle der Kombinationsmöglichkeiten in F2:
Rückkreuzung, Testkreuzung
Die Testkreuzung wendet man dann an, wenn man bei der Zucht homozygote und heterozygote Individuen trennen will. Wenn man bei dieser Kreuzung einen Elternteil nimmt, wird der ganze Prozeß als Rückkreuzung bezeichnet und die neue Generation wird mit dem Buchstaben Bgekennzeichnet (nach dem englischen Wort "backcross"). Am besten verdeutlichen wir die ganze Situation wieder an einem Beispiel: Die Farbe Schwarz ist gegenüber Braun dominant (mit leberbrauner Nase). Wenn wir also einen schwarzen Rüden BB mit einer braunen Hündin bb verpaaren, bekommen wir eine völlig schwarze F1-Generation mit Genotyp Bb. In der F2-Generation kommt es zur Entwicklung der Genotypen BB, 2x Bb, bb, aus der die Individuen BB und Bb wieder schwarz sein werden, aber Bb werden wieder Träger des erwünschten braunen Gens sein. Wenn wir die Tiere in Träger (heterozygot) und Nichtträger (homozygot) trennen wollen, müssen wir die Testkreuzung mit einem braunen Individuum anwenden, also mit einem rezessiv-homozygoten bb. Ein homozygot schwarzer Partner kann nur schwarze Nachkommen ergeben, wogegen ein heterozygot schwarzer auch braune Nachkommen haben kann.
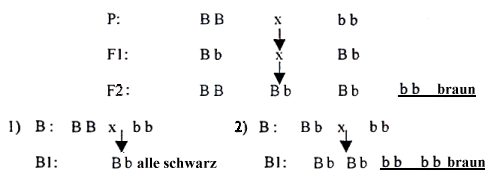
Wir dürfen aber niemals vergessen, daß die hier angeführten Berechnungen der Nachkommen Idealfälle sind, Theorie. In der Realität bekommen wir ähnliche Ergebnisse nur bei einer insgesamt großen Zahl von Versuchen – in einem Wurf kann die Relation der Genotypen ganz anders sein als das, was wir Ihnen hier vorgeführt haben. Es kann zum Beispiel passieren, daß wir den erwünschten braunen Nachkommen nur einmal bekommen oder daß umgekehrt der ganze Wurf braun ist.
Pioniere der Genetik
 Außer G. Mendel haben W. Bateson und der amerikanische Professor T. H. Morgan in der Genetik Geschichte geschrieben.
Außer G. Mendel haben W. Bateson und der amerikanische Professor T. H. Morgan in der Genetik Geschichte geschrieben.
Bateson knüpfte an Mendels Entdeckungen an und beobachtete weiter das Verhalten der Gene in der zweiten und dritten Filialgeneration. Er wurde schnell durch die Entdeckung überrascht, daß sich die Gene bei bestimmten Eigenschaften anders als erwartet verhielten, daß das Teilungsverhältnis nicht den Mendelschen Regeln entsprach und daß schließlich einige Kombinationen bei der Nachkommenschaft überhaupt nicht vorkamen.
 Aber erst Thomas H. Morgan hat erkannt, daß das Phänomen, welches er die Kopplung der Erbanlage nannte, auf die Struktur der Chromosomen und die Einbettung der Gene in ihnen zurückzuführen ist. Er entdeckte und wies nach, daß die Gene in den Chromosomen nacheinander an einem relativ festen Platz linear aufgereiht sind, den er als Locus (Genort) bezeichnete, und daß sie sich gegenseitig beeinflussen. Die gegenseitige Beeinflussung der Gene (Kopplung der Erbanlage) ist um so größer, je näher die Gene in den Chromosomen beieinander liegen. Um die Situation noch komplizierter zu machen, entdeckte Morgan auch die Tatsache, daß sich die Chromosomen gern und häufig gegenseitig überkreuzen, abreißen und wieder so verschmelzen, daß sie ihre Plätze tauschen. Dieser Wechsel in einem Chromosom kann auch mehrmals vorkommen. Auf diese Weise ändern sich der Inhalt und auch die Reihenfolge der Gene im Chromosom. Je näher die Loci beieinander liegen, um so geringer ist die Wahrscheinlichkeit, daß es zu einer Überkreuzung zwischen ihnen kommt. Im Gegenteil, je weiter sie auseinander liegen, um so wahrscheinlicher ist eine Überkreuzung.
Aber erst Thomas H. Morgan hat erkannt, daß das Phänomen, welches er die Kopplung der Erbanlage nannte, auf die Struktur der Chromosomen und die Einbettung der Gene in ihnen zurückzuführen ist. Er entdeckte und wies nach, daß die Gene in den Chromosomen nacheinander an einem relativ festen Platz linear aufgereiht sind, den er als Locus (Genort) bezeichnete, und daß sie sich gegenseitig beeinflussen. Die gegenseitige Beeinflussung der Gene (Kopplung der Erbanlage) ist um so größer, je näher die Gene in den Chromosomen beieinander liegen. Um die Situation noch komplizierter zu machen, entdeckte Morgan auch die Tatsache, daß sich die Chromosomen gern und häufig gegenseitig überkreuzen, abreißen und wieder so verschmelzen, daß sie ihre Plätze tauschen. Dieser Wechsel in einem Chromosom kann auch mehrmals vorkommen. Auf diese Weise ändern sich der Inhalt und auch die Reihenfolge der Gene im Chromosom. Je näher die Loci beieinander liegen, um so geringer ist die Wahrscheinlichkeit, daß es zu einer Überkreuzung zwischen ihnen kommt. Im Gegenteil, je weiter sie auseinander liegen, um so wahrscheinlicher ist eine Überkreuzung.
Eine praktische Auswirkung Kopplung der Erbanlage ist, daß sich einige Eigenschaften gemeinsam vererben – das Auftreten einer Eigenschaft ist abhängig vom Auftreten einer anderen. Das Auftreten von Taubheit bei weißgefärbten Welpen einiger Rassen und der schwere Zahnmangel bei fellosen Rassen ist wahrscheinlich auch ein Fall einer Kopplung der Erbanlage.
Vererbung des Geschlechts
Träger der unterschiedlichen Geschlechtschromosomen ist das Männchen, dessen Genotyp generell AAXY lautet. Die von ihm gebildeten Gameten sind also im Idealfall zu 50% Ax und zu 50% AY. Das Weibchen bildet zu 100% die Gameten AX, weil ihr Genotyp AAXX ist. Es ist also garantiert der Vater, der dafür verantwortlich ist, wie viele männliche und wie viele weibliche Nachkommen in einem Wurf geboren werden. Aus der Zuchtpraxis und aus Laborversuchen ist bekannt, daß Spermien mit Y-Chromosomen schwächer und empfindlicher sind als Spermien mit X-Chromosomen. Daher beruhen sämtliche Methoden, die zum Ziel haben, das Geschlechtsverhältnis bei den Nachkommen zu beeinflussen, darauf, gegen diese schwächeren Spermien vorzugehen – indem sie ihnen entweder auf ihrem Weg zu Ei behilflich sind oder umgekehrt versuchen, sie unterwegs auszuschalten.
Geschlechtsgekoppelte Vererbung – geschlechtsgebundene Vererbung
Die Geschlechtschromosomen sind auch Träger der Gene, die einige Merkmale und Eigenschaften beeinflussen. Die Eltern haben nicht die gleiche genetische Anlage (Vater XY, Mutter XX) und daher entwickeln sich bei ihnen auch Gene, die in den Geschlechtschromosomen liegen, nicht gleichermaßen häufig.
Einige Erbdefekte sind an das Geschlecht gekoppelt – Farbenblindheit (beim Menschen oft beim Mann, aber selten bei der Frau) oder die Bluterkrankheit (häufig auch beim Hund, bei Rüden). Für Zuchthunde hat allerdings größere Bedeutung die geschlechtsgebundene Vererbung. Hier sind beide Geschlechter genetisch gleichermaßen betroffen, in ihrem Phänotyp zeigt sich dies jedoch nicht auf gleiche Weise – was es in der Praxis schlimmer und bei der Zucht schwieriger macht!!
Ein klassischer Fall ist das Auftreten des Kryptorchismus (Zurückbleiben eines oder beider Hoden). Ein kryptorchider Rüde ist jedem sofort ersichtlich (wir meinen natürlich ein homozygot veranlagtes Individuum), aber eine Hündin des gleichen Genotyps ist auf keinerlei Weise auffällig. Ein kryptorchider Rüde muß sofort aus der Zucht genommen werden, aber wenn eine auf gleiche Weise homozygot kryptorchide Hündin von einem fehlerfreien Rüden gedeckt wird, bringt sie ihr ganzes Leben lang heterozygot veranlagte Welpen hervor, die fehlerfrei erscheinen, aber diesen Fehler weiterhin mit sich tragen und die Zucht verseuchen.
Erbanlagen außerhalb des Zellkerns
Bisher haben wir von zellkernbedingter Vererbung gesprochen, bei der die Gene in den Chromosomen in den Zellkernen lagern. Große Bedeutung für die Zuchtpraxis haben aber die sogenannten Erbanlagen außerhalb des Zellkerns, wo das Zytoplasma (Zelleib) die entscheidende Rolle spielt. Das Ei enthält weit mehr Zytoplasma als das kleine Spermium (bei dem es praktisch nur in den Geißeln vorhanden ist). Es ist bekannt, daß Erbfaktoren, die im Zytoplasma vorhanden sind, bei den Nachkommen eine größere Ähnlichkeit mit der Mutter fördern. Dieses Phänomen wird als maternale Vererbung bezeichnet. Ein typischer Fall von maternaler Vererbung ist die Größe der Welpen bei der Geburt. Wenn wir eine Hündin einer großen Rasse mit einem kleinen Rüden verpaaren, werden die Welpen hinsichtlich der Größe ihrer Mutter entsprechen. Eine Hündin einer kleinen Rasse bringt, verpaart mit einem Rüden einer großen Rasse, kleine Welpen zur Welt (entsprechend ihrer eigenen Größe), die erst im Laufe der Zeit größer werden.
Die Fellfärbung beim Hund
Die Farbe des Fells ist das erste, was wir beim Hund bemerken, und des öfteren entscheidet gerade sie, welchen Welpen eines Wurfs ein neuer Halter auswählt. Bei reinrassigen Hunden entscheidet der Rassestandard, welche Färbung erwünscht oder unerwünscht ist. Als Züchter sollte man sich daher dafür interessieren, auf welche Weise sich welche Farbe des Fells vererbt und wie man bei der Zucht verfährt, damit ärgerliche Fehler vermieden werden.
 Mit der Frage der Vererbung der Fellfärbung des Hundes haben sich eine Reihe von Forschern befaßt, meistens leider nicht allzu systematisch. Als den Begründer und Bahnbrecher des Studiums der Vererbung der Fellfärbung des Hundes kann man Clarence C. Little ansehen, der im Jahre 1957 eine bis heute gültige Gesamtübersicht der Gene veröffentlichte, die bei der Fellfärbung eine Rolle spielen.
Mit der Frage der Vererbung der Fellfärbung des Hundes haben sich eine Reihe von Forschern befaßt, meistens leider nicht allzu systematisch. Als den Begründer und Bahnbrecher des Studiums der Vererbung der Fellfärbung des Hundes kann man Clarence C. Little ansehen, der im Jahre 1957 eine bis heute gültige Gesamtübersicht der Gene veröffentlichte, die bei der Fellfärbung eine Rolle spielen.
Clarence C. Little war gewissermaßen der Jules Verne der Genetik: obwohl zu seiner Zeit nur wenige entscheidende Faktoren der Fellfärbung bekannt waren, ging er von ihrer Existenz aus – und spätere Forschungen haben ihm recht gegeben. Seine Kennzeichnung der einzelnen Allele ist sehr übersichtlich und logisch und daher benutzen auch wir sie.
Auch wenn es viele Hunderassen gibt, die der Mensch zielstrebig gezüchtet hat, gehören doch alle zu einer einzigen Gattung: dem Hund (canis). Es ist daher völlig logisch, wenn wir annehmen, daß für die ganze Gattung gilt, daß gleiche Farben durch dieselben Gene bedingt sind, z.B. die schwarze Farbe des Neufundländers durch dieselben Gene verursacht ist wie die schwarze Farbe des Pudels, Shar Peis oder Schipperkes. Es gibt keine Untersuchung, die dagegen
spricht, daß dem so wäre, aber selbstverständlich ist bekannt, daß manche Allele nicht bei allen Rassen vorkommen.
Bevor wir mit der ausführlichen Beschreibung der Gene, die für die Fellfärbung beim Hund relevant sind, beginnen, möchten wir eines klarstellen: wenn man von der Farbe des Hundes spricht, muß man eindeutig sein. Wenn von einem braunen Hund die Rede ist, ist immer ein Hund gemeint, der aufgrund brauner Pigmente gefärbt ist, mit einem Fell in verschiedenen Braun- oder Schokoladeschattierungen und mit einen braunen (leberbraunen) Nase. Ein solcher Hund hat an seinem ganzen Körper keinerlei schwarze Pigmente, man findet bei ihm kein schwarzes Haar und er hat auch hellere Augen und helle Krallen. Wenn wir von einem gelben Hund sprechen, meinen wir einen solchen, der schwarze Pigmente hat, also eine schwarze Nase und ein Fell mit verschiedenen Gelbschattierungen – von cremefarben über beige, golden, rehfarben, rot bis zu dunkel-mahagoni. Aber Vorsicht! Auch ein gelber Hund kann eine schlecht gefärbte Nase haben, so daß auf den ersten Blick der Eindruck entsteht, daß die Färbung auf braune Pigmente zurückgeht. Daher muß man auf die Farbe der Augen und Krallen achten und im Fell nach Andeutungen von Schwarz suchen, um den Hund richtig zu identifizieren!
Es sind ein Dutzend Grundgene bekannt, die die Fellfärbung beim Hund
beeinflussen.
Locus A:
Am Locus A gibt es eine Reihe von Allelen, die die Anzahl und Verteilung der dunklen (schwarzen, braunen) Pigmente bestimmen.
|
As
|
Dominantes Gen, das allen anderen Allelen übergeordnet ist, ermöglicht die Verteilung der dunklen Pigmente am ganzen Körper (schwarze Dogge, brauner Pudel).
|
|
ay
|
Schränkt das Auftreten der dunklen Pigmente ein und sorgt für eine gelbe Färbung in verschiedenen Tönungen (Akita, Shiba, gelbe Dogge). Die Welpen kommen mehr dunkel zur Welt, oft mit einem schwarzen Streifen auf dem Rücken. Einige Autoren sehen dieses Allel ebenfalls als dominant an, also
Ay
– aber das ist unlogisch. Wenn am Locus A zwei dominante Allele wären, obendrein für ganz unterschiedliche Farben
(As
für Schwarz und
Ay
für Gelb), wie würde dann das Ergebnis einer Verpaarung eines schwarzen Hundes mit einem gelben aussehen? Zuchtversuche beweisen ganz deutlich, daß die gelbe Farbe gegenüber der reinschwarzen Farbe rezessiv ist.
|
|
aw
|
Elementare, wolfsgraue Färbung (Deutscher Schäferhund, Wolfsspitz).
|
|
as
|
Sattelfärbung – teilweise dominant über at, erzeugt eine Doppelfärbung (bicolour) – dunkel (schwarz, braun) mit gelben Abzeichen, wobei die dunkle Farbe allein auf den Rückensattel beschränkt ist. Der ganze Kopf und der Rest des Körpers ist gelb in verschiedenen Schattierungen (Airedale Terrier, Welsh Terrier).
|
|
at
|
Ermöglicht eine Bicolour-Färbung – dunkel und gelb – derart, daß der größte Teil des Körpers dunkel ist und Gelb am Kopf punktuell über den Augen und rund um den Fang möglich ist (Dobermann, Dackel, Black-and-tan-Shiba). Zuchtversuche bei einigen Rassen zeigen, daß beim Allel
at
bestimmte Modifikatoren wirken (Polygene ?), die eine sog. Grizzly-Färbung beim Saluki oder Domino beim Afghanen hervorrufen. Es ist nicht ausgeschlossen, daß auch die typische Gesichtsfärbung beim Alaska Malamut und beim Sibirischen Huskie ähnlichen Ursprungs ist.
|
|
a
|
Rezessives Allel, beschrieben beim Deutschen Schäferhund und beim Norwegischen Elchhund, erzeugt im Verbund mit dem Gen B (also aaBB) schwarze Exemplare, rezessiv gegenüber jeder anderen Farbe.
|
Locus B:
|
B
|
Dominantes Allel, das die Bildung schwarzer Pigmente am Körper des Hundes
erlaubt.
|
|
b
|
Rezessives Allel, das die Bildung schwarzer Pigmente verhindert und im homozygoten Zustand bb braune Pigmente und eine braun gefärbte Nase und Fell hervorruft.
|
Locus C:
Am Locus C sind Allele zu finden, die vor allem die Intensität der Farbe Gelb
beeinflussen. Die Farbe Gelb hat neben der Scheckung die größte Variabilität, von optisch fast weiß bis dunkel-mahagoni (Irish Setter). Die wechselseitigen Beziehungen der Allele an diesem Locus und die Ursache weiterer Faktoren haben viele Forscher beschäftigt. Aus ihrer Arbeit und aus der Zuchtpraxis stammen die folgenden Ausführungen.
|
C
|
Ermöglicht eine satte Färbung vor allem der Farbe Gelb.
|
|
cch
|
Beeinflußt das gelbe Pigment und hellt es optisch fast bis zu weiß auf (silbergrauer Schnauzer, Kuvasz). Von der Existenz dieses Allels sind viele Forscher nicht überzeugt und neueste Forschungen zeigen, daß es sehr wahrscheinlich ist, daß anstelle eines einzelnen Gens eine Gruppe von Polygenen die Aufhellung der Farbe Gelb beeinflußt. Aus Gründen der Übersichtlichkeit werden wir aber im weiteren Verlauf diese Kennzeichnung von Little verwenden, gleichgültig ob der Grund der Aufhellung einfacher Natur ist (ein einzelnes Allel cch)
oder komplexer (eine Gruppe von Polygenen).
|
|
ce
|
Bewirkt eine außergewöhnliche Verdünnung der Farbe Gelb und sehr helle
Raubtieraugen.
|
|
ca
|
Anlage zum totalen Albinismus, wobei auf dem Körper des Hundes keinerlei
Pigmente vorhanden und die Augen rot sind.
|
Außer den besagten Allelen, die Forscher am Locus C beschrieben und Züchter nachgewiesen haben, gibt es noch eine Reihe weiterer Faktoren, die an der Gelbfärbung des Hundefells beteiligt sind. Vor allem sind es die schon erwähnten Polygene, genannt Rufus-Polygene, die die Aufhellung von Gelb beeinflussen. Aus der Praxis ist bekannt, daß die Gelbtönung stark variiert und daß es schwierig ist, die gewünschte Schattierung zu erzielen. Die Forschungsergebnisse weichen so sehr von einander ab, daß man schlecht eindeutig sagen kann, wie groß die Wechselwirkung aller Faktoren in Wirklichkeit tatsächlich ist. In Anbetracht dessen, daß nicht bekannt ist, daß zwei hellere Tiere stark gelbe Nachkommen erzeugt hätten, gehen wir davon aus, daß Littles Kennzeichnung der für die Aufhellung von Gelb zuständigen Gene als rezessiv zutreffend ist. Die Rufus-Gene beeinflussen auch den Sättigungsgrad von Schwarz – wir vermuten ihren Einfluß bei der Farbminderung des Sattels beim Airedale, beim Lakeland Terrier oder bei der Entstehung von Mattschwarz (optisch Schwarzbraun) beim Shar Pei.
Eine Gruppe von Polygenen, die Umbrous-Polygene genannt werden, erlaubt die Bildung sog. Zobelfärbungen (Sable, Sesam, Zobel, Chinchilla). Diese Gruppe beeinflußt ay und bewirkt die Vermischung gelber mit schwarzen Haaren oder die Schwarzfärbung gelber Haarspitzen. Diese Färbung ist sehr wirkungsvoll z.B. beim Saluki oder beim Tibetanischen Spaniel, dagegen äußerst unerwünscht beim Shar Pei. Es ist züchterisch erwiesen, daß es noch weitere Modifikatoren geben muß, die am Locus C wirksam sind. Es handelt sich um den Verursacher der sog. Urajiro-Färbung, der z.B. beim Saluki bekannt ist und beim Akita und Shiba direkt vom Standard verlangt wird. Genau genommen handelt es sich um eine Gelb-Variante der bereits erwähnten Grizzly-Färbung beim Windhund. Tiere mit Urajiro haben über den Augen, um den Fang, an der Kehle und Brust, an der Unterseite der Rute und Läufe eine sehr helle, optisch bis zum Weiß gehende Färbung. Zuchtergebnisse zeigen, daß es sich um rezessive Faktoren handelt, aber leider hat sich mit dieser Erbanlage bisher niemand beschäftigt.
Locus D:
|
D
|
Dominantes Allel, beteiligt an der Bildung schwarzer Pigmente.
|
|
d
|
Rezessives Allel, das das schwarze Pigment nach Blau wandelt. Die Welpen werden bereits blau geboren (blaue Dogge)! Hunde mit diesem Gen haben keine schwarze Nase, vielmehr ist sie schiefergrau (sofern das Gen in der homozygoten Form dd vorliegt!) und ihre Augen sind gleichfalls heller. Dieses Allel beeinflußt selbstverständlich das schwarze Pigment auch bei gelben Hunden, die dann eine schiefergraue Nase und ein Fell haben, das nicht eindeutig gelb ist, vielmehr isabell-pastell (Italienisches Windspiel, American Staffordshire Terrier). In Zusammenarbeit mit dem Gen b (in der Form bb) ruft es eine silbergraue Färbung hervor, bekannt beim Weimaraner und beim Border Collie.
|
Locus E:
|
E
|
Erlaubt die Verteilung dunkler Pigmente im ganzen Fell.
|
|
e
|
Ermöglicht die Bildung dunkler Pigmente im Fell und wandelt in der homozygoten Form ee Schwarz nach Zitronengelb. Die Welpen werden eindeutig gelb geboren, ohne Andeutung schwarzer Haare am Körper. Diese Färbung ist allgemein äußerst unerwünscht, weil sie bei der Zucht viele Probleme verursacht. Sie ist beim Dalmatiner bekannt (gelbe Punkte, aber eine schwarze Nase) oder beim "unechten" Aprikot-Pudel, der ein gelbes Fell, aber ebenfalls eine schwarze Nase hat.
|
|
ebr
|
Beeinflußt die Stromung des Fells. Experimente haben gezeigt, daß die Veranlagung zur Stromung tatsächlich rezessiv ist, auch wenn es Autoren gibt, die behaupten, daß es sich um ein dominantes Gen handelt. Zu dieser Fehleinschätzung sind sie wahrscheinlich durch die große Variabilität der Stromung gekommen. Es sind Einzeltiere bekannt, die schwarz ausschauen, aber bei näherer Ansicht merkt man, daß sie einige hellere Haare am Körper haben, die entfernt eine Stromung andeuten (Französische Bulldogge, Staffordshire Bullterrier). Wiederum gibt es auch solche Exemplare, bei denen die Stromung nur auf der Haut offensichtlich ist und im Fell nur im zarten Alter. Im Laufe der Zeit verschwindet die Stromung aus dem Fell, das Fell ist rein gelb und versuchsweise ist ein solches Tier fälschlich als gelb ohne Stromung angesehen worden (Whippet, Irischer Wolfshund). Die Intensität der Stromung wird wiederum durch eine Reihe von Polygenen beeinflußt, deren genaue Verbindung bisher nicht eingehend untersucht worden ist.
|
Am Locus E ist auch die Existenz des Allels Em
beschrieben, das allen anderen Alleln übergeordnet ist, welche die Bildung der Maske (schwarz, braun) beeinflussen. Einige Autoren führen an, daß bei Gelbfärbung auf der Grundlage ee sich keine Maske entwickelt und daß das eine sichere Art ist, um ein "echt" gelbes Tier im Unterschied von einem unerwünschten zitronengelben zu unterscheiden.
Locus G:
|
G
|
Teilweise dominantes Gen, das in Zusammenarbeit mit anderen Genen dunkel (schwarz, braun) geborene Welpen hell (grau, helles kakaobraun) werden läßt. Typische Beispiele sind der Kerry Blue Terrier und der Cesky Terrier.
|
|
g
|
Rezessives Allel, das diese Farbaufhellung nicht bewirkt.
|
Locus M:
|
M
|
Dominantes Gen, das die Bildung der sog. Merle-Zeichnung beim Collie, Dackel, Harlekin-Dogge und weiteren Rassen bewirkt. Ein Hund mit diesem Faktor hat am Körper außer dunklen Flecken (schwarz, braun) auch helle Flecken (grau, kakaobraun). Falls die Augenpartie hell ausfällt, ist auch ein Pigmentmangel in der Regenbogenhaut (Iris) des Auges wirksam. Dieses Gen ist letal – Welpen mit MM werden tot geboren oder gehen schnell zugrunde. Daher werden in der Zucht zwei Merle-Tiere besser nicht verpaart.
|
|
m
|
Rezessives Allel, das eine normale Färbung erlaubt.
|
Locus P:
|
P
|
Behält die Farbsättigung bei.
|
|
p
|
Rezessives Allel, das sehr selten vorkommt. Es wurde vor allem beim Pekinesen beschrieben, bei dem das schwarze Fell-Pigmente rauchfarben und dunkle Augen rosa werden.
|
Locus R:
|
R
|
Dominantes Gen, das die sog. Roan-Färbung beeinflußt, bei der ein Haar weiß und eins pigmentiert ist. Wurde beim Cocker Spaniel entdeckt und beschrieben.
|
|
r
|
Bewirkt keine Änderung.
|
Locus S:
|
S
|
Dominantes Gen, das eine dunkle Fellfärbung am ganzen Körper erlaubt. Wenn unter dem Einfluß dieses Gens weiße Flecken entstehen, ist das nur eine zufällige Entwicklung auf einer kleinen, weißen Fläche, ausgelöst durch den sog. spontanen bzw. zufälligen Albinismus. Zu einem solchen Verlust der Pigmente bei ganzfarbigen Rassen kommt es öfters an der Brust und an den Zehen der Vorderläufe.
|
|
si
|
Rezessives Allel, das die sog. "Irische Scheckung" erlaubt, die für einige Rassen typisch ist (Collie, Bullterrier). Ein Hund mit diesem Gen in homozygoter Form hat eine weiße Blesse am Kopf, einen weißen Halskragen unterschiedlicher Breite, weiße Pfoten und eine Rute mit weißer Spitze.
|
|
sp
|
Rezessives Allel, das eine größere Bildung weißer Flecken hervorruft, die sog. echte Scheckung. Das Ausmaß der farbigen Fläche reicht von 80% bis 20% der Gesamtfläche des Körpers.
|
|
sw
|
Niedrigstes Allel, das eine noch größere Fläche weißer Flecken erlaubt. Die farbige Fläche beträgt dann weniger als 20% der Körperoberfläche oder die Pigmente beschränken sich auf die Augenlider und den Rand der Lefzen (Sealyham Terrier).
|
Alle Allele an diesem Locus überlappen einander in der Wirkung und rufen bei heterozygoter Kombination unterschiedlich große Mengen weißer Abzeichen hervor. Zum Beispiel hat ein Tier mit
si si
weniger Weiß als eines mit
si sp
oder sogar eines mit
si sw
usw.
Locus T:
Beeinflußt die Reinheit der weißen Fläche.
|
T
|
Dominantes Allel, das die Sprenkelung/Tüpfelung der weißen Fläche beeinflußt.
|
|
t
|
Rezessives Allel, das die Reinheit der weißen Fläche beibehält.
|
So wie einige Standards eine rein weiße Fläche verlangen (King Charles Spaniel), fordern andere umgekehrt eine gesprenkelte Fäche (Bohemian Spotted Dog, Dalmatiner). Beim Dalmatiner wurde lange angenommen, daß es sich um ein besonderes Gen handelt, das die typische Musterung hervorruft. Heute weiß man, daß es sich tatsächlich um eine klassische Sprenkelung (Ticking) handelt (T), die die Züchter durch die Auswahl von Eltern-Paaren zu einer solchen Vollendung gebracht haben.
Locus W:
An diesem Locus liegen Allele für die dominante Weißfärbung, die sich nur bei einigen Rassen entwickeln. Forscher konnten diesen Faktor dank des Bullterriers nachweisen, bei dem Weiß über Färbung dominant ist.
|
W
|
Beeinflußt die Weißfärbung des Fells beim Bullterrier.
|
|
w
|
Behält die Fellfärbung bei.
|
Fellfärbung in der Zuchtpraxis
Wir haben ausgeführt, wie die Gene die Fellfärbung des Hundes beeinflussen. Wir wissen, daß Großbuchstaben dominante Gene bezeichnen, die die Bildung dominierender, auf den ersten Blick auffälliger Eigenschaften bewirken, während Kleinbuchstaben rezessive Gene bezeichnen, die unterdrückte Eigenschaften steuern, welche vielleicht schon seit Generationen verdeckt sind.
Wie können wir diese Erkenntnisse praktisch anwenden?
Zum Beispiel so:
Es gibt bei den Hunden eine unabänderliche Reihenfolge der grundsätzlichen Färbungen je nachdem, welche Allele des Gens A sie haben. An oberster Stelle ist die vollständig schwarze Färbung (Neufundländer, Dogge ...) – aber Vorsicht bei einigen Ausnahmen, darunter Deutsche Schäferhunde und Elchhunde! Ein schwarzer Hund kann Anlagen zu allen anderen Fellfärbungen in sich tragen und nur in der Zucht, d.h. durch Versuche bei der Verpaarung, kann man ans Licht bringen, welche das sind. Ein schwarzes Paar kann schwarze Welpen haben, gelbe, wolfsgraue und zweifarbige.
Der Shiba hat kein Gen für eine vollständig schwarze Färbung. Unter den verwandten Rassen ist nur der Kai schwarz.
Auf Schwarz folgt Gelb in allen Abstufungen und Modifikationen. Dazu zählen das Mahagoni der Irish Setter, desgleichen das Goldgelb und Sesam der Japaner, das Sandgelb der Doggen, der Windhunde und Terrier oder das Cremefarbene des Hokkaido, des Kishu, des Akitas, des Slovensky Cuvac usw. Zwei gelbe Einzeltiere mit dem Gen ay können zusammen niemals vollständig schwarze Welpen haben (ich erinnere nochmals an die seltene Ausnahme beim Deutschen Schäferhund), sie können jedoch wolfsgraue und zweifarbige Kinder haben. Dies ist auch beim Shiba so: gelbe Exemplare haben gelbe Welpen in verschiedenen Abstufungen, sesam (Einfluß der Umbrous-Polygene auf das Gen ay bei der Farbe Gelb) oder schwarz mit gelben Zeichen. Das Gen für die Wolfsfärbung aw hat der Shiba nicht, aber von den verwandten Rassen ist es typisch für den Shikoku. Ob ein Shiba eine Anlage zu schwarz-gelber Färbung hat oder nicht hat, kann man immer nur experimentell, durch Zuchtversuche ermitteln. Weil aber die Anlage zur Bicolour-Färbung at gegenüber der Anlage Gelb ay rezessiv ist, kann es einige Generationen dauern, bevor sie in der Zucht zu Tage tritt. Wir kennen dies auch aus unserer eigenen Zucht: die erste ins Zuchtbuch eingetragene Shiba-Hündin Manlötens Yubina war Träger des Gens at, aber erst ihr Nachkomme, die Hündin Ichidó, brachte mit dem Rüden Mara-Shima Timo (gleichfalls Träger des Gens at) schwarz-gelbe (black and tan) Welpen zur Welt.
Zwei wolfsgraue Tiere können zusammen nur wolfsgraue oder zweifarbige Welpen haben, weil sie in der Hierarchie der Hundefarben an dritter Stelle stehen, erst nach vollständig schwarz und gelb.
Ganz unten stehen die sog. Doppelfärbungen, also schwarz (braun, blau) mit gelben Zeichen. Zwei schwarz-gelbe Einzeltiere können zusammen niemals einfarbige Welpen haben, nur zweifarbige. Wenn es vorkommt, daß sie z.B. rote Kinder
haben, ist es ein Beleg, daß sie nicht beide die Eltern des Wurfs sein können!
Alle Färbungen können mit weißen Zeichen einhergehen oder können eine Kombination mit einer größeren Menge Weiß sein, d.h. sie können gefleckt sein.
Als Fleckung in genauen Sinn des Wortes kann man die Bildung größerer weißer, nicht-pigmentierter Flächen auf einem ansonsten pigmentierten Körper, Kopf und Gangwerk ansehen. Gefleckt sind also nicht sog. zweifarbige Hunde (schwarz, braun, blau mit gelben Abzeichen). Auch nicht solche Rassen, bei denen ein gelber Bereich so stark aufgehellt ist, daß er optisch weiß wirkt, z.B. bei Alaskan Malamut, Siberian Husky oder Shiba. Eine nicht-pigmentierte Behaarung ist auch bei pigmentierten Rassen im Brustbereich oder an den Zehen der Vorderläufe zu finden. Bei einigen Rassen werden diese kleinen weißen Flecken als Fehler angesehen, bei anderen erlaubt sie der Standard. Sehr große weiße Abzeichen, größer als nur ein paar weiße Haare im Brustbereich oder an den Zehen der Vorderläufe, werden als spontaner Albinismus bezeichnet. Dabei handelt es sich um eine völlig zufällig entstandene Depigmentierung (Pigmentschwund). Es ist aber erwiesen, daß der Umfang nicht-pigmentierter Flächen sich zu vergrößern beginnt, sobald man Partner mit weißen Flecken paart. Das Weiß verbreitet sich auf dem Körper des Hundes folgendermaßen: es beginnt mit kleinen weißen Flecken an der Brust und an den Vorderpfoten. Dann werden das Kinn und die ganzen Vorderpfoten weiß. Später breitet sich das Weiß auf die Zehen der Hinterläufe aus, und auch die Spitze der Rute ist weiß. Schließlich greifen die Zeichen von der Brust auf den Kopf über sowie auf den hinteren Teil des Körpers bis zum Bauch. Die Flecken am Kinn schreiten zum Oberkiefer fort, wo eine Blesse unterschiedlicher Größe entsteht. Das Weiß erstreckt sich vom Hals bis zur Unterseite des Halses – zuerst als Fleck im Nacken, später als unterschiedlich breiter Halskragen. An den Läufen steigt das Weiß auf in Richtung Rückgrat. Dieses Stadium der Verbreitung der weißen Farbe – Blessen am Kopf und Hals, ganz weiße Läufe und eine weiße Rutenspitze – bezeichnet man als "Irische Fleckung" und man weiß, daß sie durch das rezessive Gen si beeinflußt wird. Eine solche Färbung kann man züchterisch ganz gut erreichen, und es gibt Rassen, bei denen sie nicht nur oft auftritt (Bullterrier, American Staffordshire Terrier), sondern wo sie der Standard sogar vorschreibt (Collie). Homozygote Tiere mit si si haben alle ungefähr gleich große Abzeichen, und durch Auswahl der Zuchtpaare kann man Abweichungen gut eliminieren.
Von der Irischen Fleckung aus schreitet die Depigmentierung sehr rasch voran. Der weiße Halskragen weitet sich so sehr aus, daß er den ganzen Hals vom Schulterblatt bis zum Widerrist umfaßt und den pigmentierten Kopf vom Körper trennt. Das Weiß an Brust und Bauch steigt bis zur Höhe der Ellbögen, ebenso das Weiß an den Läufen. Am Körper ist nur noch die obere Bedeckung farbig, pigmentiertes Fell bleibt am Kopf, hauptsächlich an den Ohren und um die Augen herum. Im nächsten Stadium verbindet sich das Weiß an den Leisten mit dem Rückgrat – die Decke zerreißt in einzelne Flecken. Die Blesse am Kopf verlängert sich, sie verbindet sich mit dem Halskragen und teilt die rechte und linke Seite des Kopfes in zwei selbständig pigmentierte Teile. Die weiße Farbe an der Rute nimmt derart zu, daß fast die ganze Rute weiß ist. Der Hund ist gefleckt, und es ist erwiesen, daß er Träger des Allels sp ist, welches diese Färbung (die sog. "echte Fleckung") hervorruft. Dieses Allel ist rezessiv gegenüber dem Allel si für die Irische Fleckung. Heterozygote Hunde mit si sp sind weniger gefleckt als homozygote mit sp sp.
Es gibt drei Stellen beim Hund, wo sich pigmentiertes Fell am längsten hält: der Kopf um die Augen, die Mitte des Rückens und die Wurzel der Rute. Bei fortschreitender Weißwerdung schwindet zuerst die Pigmentierung am Rücken, dann die Färbung an der Wurzel der Rute. Am längsten hält sich die Pigmentierung am Kopf, meistens verliert sich eher die Farbe um die Augen und hält sich länger an den Ohren. Tiere, die Pigmentierungen nur am Kopf haben oder schließlich bis auf Augen, Nase und Lefzen ganz weiß sind, bezeichnet man als extrem gefleckt, genetisch sw sw. Das Alell sw rangiert an unterster Stelle am Locus S und ruft den größten Verlust pigmentierter Fläche am Körper des Hundes hervor. Heterozygote Hunde mit si sw und sp sw sind immer mehr pigmentiert als homozygote mit sw sw.
Weiße Flächen sind entweder rein weiß oder gesprenkelt/getüpfelt (infolge des Allels T). Die Punkte können klein oder groß sein, dünn oder dicht verteilt. Manchmal sind sie so dicht, daß es bei einem langen Fell den Anschein hat, als wenn der Hund gestromt (gestreift) wäre oder sogar roan (geschimmelt: ein Haar weiß, eins pigmentiert). Die Farbe der Punkte oder Flecken wird durch die Lage am Körper des Hundes und durch die Erbanlage bestimmt. Jeder gefleckte Hund hat unter seiner weißen Färbung seine ursprüngliche Farbe verborgen – wenn er nicht weiß oder gefleckt wäre, wäre er eigentlich schwarz, gelb, gestromt, schwarz mit gelben Zeichen usw. Die Farbe der Punkte ist abhängig von der Ursprungsfarbe: Punkte (Flecken) auf einer schwarzen Fläche sind schwarz, Punkte in der Nähe von Gelb sind gelb. Oder umgekehrt, aus der Farbe der Punkte (Flecken) am Körper des Hundes kann man auf die zugrundeliegende Farbe schließen: schwarze Punkte am Rücken und gelbe an den Beinen deuten darauf hin, daß der Hund eigentlich schwarz mit gelben Zeichen ist, schwarze und gelbe Punkte vermischt am ganzen Körper und an den Beinen zeigen, daß der Hund eigentlich gestromt ist usw.
Ein besonderes Beispiel einer Kombination von extremer Fleckung und weiß gepunkteter Flächen ist der Dalmatiner. Daß er Träger des Gens sw ist, verrät sich dadurch, daß beim ihm häufig unerwünscht dunkle Flecken, die größer als Punkte sind, auftreten. Die charakteristische Sprenkelung/Tüpfelung ist also eine Folge langjähriger Zuchtpraxis, durch die sowohl die Form und Größe als auch die Häufigkeit der Punkte standardisiert wurden. Beim Shiba sind große weiße Abzeichen unerwünscht und Fleckung ist ein Ausschlußfehler. Nichtsdestotrotz sind z.B. in den USA gefleckte Shibas sehr beliebt und gesucht.
Die Fellfärbung beim Shiba
Der Shiba-Standard erlaubt drei Färbungen: rot/gelb (*), sesam und schwarz mit gelben Abzeichen. Alle Färbungen müssen urajiro haben, also ein helleres Fell um den Fang herum, an der Kehle, am Unterkörper, an der Rute und an den Läufen. Einige Shibas haben außer urajiro auch weiße Abzeichen unterschiedlicher Größe, öfters an den Pfoten und an der Brust. Es gibt keine völlig schwarzen Shibas, auch keine wolfsgrauen, gestromten, blauen oder schokoladebraunen.
[* Anm. des Übersetzers: In Tschechien werden für die drei Standardfarben in zwei Fällen andere Bezeichnungen verwendet als international üblich. Daher sind ein paar Bemerkungen zur Namensgebung unvermeidbar.
Die Farbe "sesam" wird auch tschechisch so bezeichnet. Statt "black and tan" wird meistens "schwarz mit gelben Abzeichen" bzw. "schwarz-gelb" oder auch "schwarz mit Brand" verwendet. Richtigen Diskussionsstoff bietet dagegen die dritte Fellfarbe.
International wird stets "rot" (red) als zulässige Fellfarbe des Shibas angegeben. Dies ist aus genetischer Sicht fragwürdig, da das "Rot" der Shibas durch Allele am Locus C, die vor allem die Intensität der Farbe Gelb beeinflussen, zustande kommt (mehr dazu unten). Man muß dazu bedenken, daß die heute gültige englische Übersetzung des Shiba-Standards von 1987 auf den späteren japanischen Nihon Ken Hozonkai-Standard und nicht auf den ursprünglichen Nippo-Standard von 1935 beruht, der 9 verschiedene Farbschattierungen (tawn, white, wheaten usw.) für den Shiba nennt, darunter aber nicht rot. Offensichtlich ist das "red" des internationalen Standards nur ein Sammelbegriff für die verschiedenen Farbtöne des ursprünglichen japanischen Standards. In Tschechien wird daher meistens das Wort "zlatá" (= goldfarben) oder auch "žlutá" (= gelb) für die Fellfarbe des Shibas verwendet, wenn nach unserem Verständnis "rot" gemeint ist. Als Kompromiß haben wir "zlatá" im Folgenden mit "goldrot" übersetzt, "žlutá" dagegen bei "gelb" belassen.–
Diese Konfusion bei der Benennung der Farben ist im übrigen nichts Shiba-spezifisches: so wird z.B. im englischen Standard für den Dobermann-Pinscher die braune Fellfarbe als "red" bezeichnet, während sie in der deutschen Version des Standards auch als braun angegeben wird.]
Schauen wir einmal, welche Gene die Fellfärbung beim Shiba beeinflussen.
1. Gelb in allen Abstufungen
Shibas werden in einer großen Bandbreite von Gelbfärbungen gezüchtet. Die hellste Färbung ist fast weiß, sahne- oder cremefarben, nur mit einer dunklen Schattierung an den Ohren. Eine solche Färbung hatte die Hündin Orion's Minimeadow Sugar, ein USA-Import, die in den Stammbäumen vieler tschechischer Shibas auftaucht.
 Ganz dunkle, wirklich "rote" Shibas sind sehr selten, sie haben ein satt mahagonifarbenes Fell. Wenn sie ein ausgeprägtes Urajiro haben, sind sie sehr beliebt und in Japan äußerst begehrt. Eine solche Färbung hat der Rüde Shingen Go Chousei Sou (siehe Bild), ein Import aus Japan, der auf der Spezial-Ausstellung des SHIBA KLUBS 2003 in Zbraslav von seinem holländischen Halter vorgeführt wurde. Die Mehrzahl der Shibas liegt zwischen diesen beiden Extremen, sie haben ein mittleres Gelb, goldrot eingefärbt.
Ganz dunkle, wirklich "rote" Shibas sind sehr selten, sie haben ein satt mahagonifarbenes Fell. Wenn sie ein ausgeprägtes Urajiro haben, sind sie sehr beliebt und in Japan äußerst begehrt. Eine solche Färbung hat der Rüde Shingen Go Chousei Sou (siehe Bild), ein Import aus Japan, der auf der Spezial-Ausstellung des SHIBA KLUBS 2003 in Zbraslav von seinem holländischen Halter vorgeführt wurde. Die Mehrzahl der Shibas liegt zwischen diesen beiden Extremen, sie haben ein mittleres Gelb, goldrot eingefärbt.
Die Intensität der Farbe Gelb wird außer durch eigene Gene am Locus C auch durch verschiedene rezessive Polygene (eine Gruppe kleinerer, schwächerer Gene, die nicht selbständig funktionieren, sondern immer nur in der Gruppe) verursacht, vor allem durch die Rufus-Polygene, die Gelb aufhellen.
Aus der Praxis ist bekannt, daß die Intensität von Gelb stark variiert und daß es sehr schwierig ist, den gewünschten Farbton zu erzielen. Es ist aber auch erwiesen, daß zwei helle Tiere zusammen keine satt gelben Nachkommen haben, wodurch die Theorie des rezessiven Ursprungs des Polygens für die Gelbaufhellung untermauert wird.
Das Sesam gelbgefärbter Exemplare kommt unter dem Einfluß der Umbrous-Polygene zustande. Diese bewirken eine Durchmischung des gelben Fells mit schwarzen Haaren oder nur die Schwarzfärbung der gelben Haarspitzen. Man kann aber auch bei dieser Färbung nicht abschätzen, wie intensiv die Sesamfarbe eines Hundes ausfallen wird. Je stärker bei den Eltern die Durchmischung des schwarzen Fells mit Gelb ist, desto wahrscheinlicher ist es, daß ihre Nachkommen ein dunkles Sesam haben werden. Leider ist es aber des öfteren ein sog. unreines Goldrot, bei dem in dem gelben Fell nur ein paar schwarze Haare sind, größtenteils an der Rute und am Widerrist.
Völlig unerforscht sind die Modifikatoren, die für die Bildung von Urajiro sorgen. Die Zuchtpraxis hat erwiesen, daß es sich sehr wahrscheinlich um rezessive Polygene handelt, die die Entstehung, das Ausmaß und den Helligkeitsgrad an bestimmten Körperpartien beeinflussen. Bisher haben sich die Wissenschaftler nicht näher mit dieser Problematik beschäftigt. Es steht aber fest, daß ein großes und sehr helles Urajiro beim Shiba ebenso schwer zu erreichen ist wie eine Sesamfärbung am Großteil des Körpers. Goldrote und sesamfarbene Shibas besitzen u.a. die folgenden Gene für ihre Fellfärbung: ay B C cch (entweder ein einzelnes Gen oder eine Gruppe von Polygenen).
Schwarz mit gelben Abzeichen (black and tan)
Eine sogenannte Doppelfärbung, beim Shiba ausschließlich schwarz (niemals blau oder schokoladenfarben) mit gelben Abzeichen über den Augen, um den Fang, an der Kehle, am Bauch und an der Unterseite der Läufe, ist in der Farbhierarchie des Hundes eine ganz unten angesiedelte Färbung, weil sie durch das Gen at zustande kommt, dem letzten aller Allele des A-Gens. Ein zweifarbiges Exemplar kann von gelben Eltern abstammen, sofern diese selber die Anlage at besitzen, aber untereinander können schwarz-gelbe Partner nur schwarz-gelbe Nachkommen haben.
 Auch ein schwarz-gelber Shiba muß ein markantes Urajiro haben, das normalerweise nicht so hell ist wie bei gelben Exemplaren. Bei einem dunklen Shiba ist eine große helle Hemdbrust (Chemisette) erwünscht, die die helle Flächen um den Fang mit dem hellen Fleck an der Brust verbindet (siehe Bild). Der Umfang des Urajiro ist auch bei schwarz-gelben Shibas sehr zufallsabhängig, und der gewünschte Effekt ist nur sehr schwer zu erreichen. Das wissen die Japaner ganz genau und daher siegen schwarz-gelbe Shibas mit großem Urajiro auf den prestigeträchtigen Ausstellungen zu Hause oft über die namhaften goldroten Champions. Zweifarbige Shibas haben also die Gene at B C cch.
Auch ein schwarz-gelber Shiba muß ein markantes Urajiro haben, das normalerweise nicht so hell ist wie bei gelben Exemplaren. Bei einem dunklen Shiba ist eine große helle Hemdbrust (Chemisette) erwünscht, die die helle Flächen um den Fang mit dem hellen Fleck an der Brust verbindet (siehe Bild). Der Umfang des Urajiro ist auch bei schwarz-gelben Shibas sehr zufallsabhängig, und der gewünschte Effekt ist nur sehr schwer zu erreichen. Das wissen die Japaner ganz genau und daher siegen schwarz-gelbe Shibas mit großem Urajiro auf den prestigeträchtigen Ausstellungen zu Hause oft über die namhaften goldroten Champions. Zweifarbige Shibas haben also die Gene at B C cch.
Das genetische Muster für die Fellfärbung beim Shiba ist folgendermaßen:
| ay |
B |
C |
D |
E |
g |
m |
P |
r |
S |
t |
w |
| at |
|
cch (oder eine Gruppe von Polygenen) |
si |
|
|
| |
|
|
|
|
|
|
|
|
sp |
|
|
|
Ing. Hana Petrusová
Zuchtberaterin SHIBA KLUB
Aus dem Tschechischen von Holger Funk.
